Triinugeneetika ja molekulaartrinoloogia
Selle peatüki tegelik eesmärk on küll tutvustada triinude üldist bioloogiat (eeskätt sigimisbioloogiat), aga kahjuks ei saa me mitte kuidagi ilma molekulaartasandile laskumata seda teha. Katsun siiski siinkohal mitte sukelduda triinude biokeemia erinevate tahkude lahkamisse ja piirduda vaid sellega mis möödapääsmatu. Alustuseks heidame pilgu peale klassikalisele triinugeneetikale.
Klassikaline triinugeneetika
Samamoodi nagu triinudest endist, nii oli ka sigimisest pikka aega väga vähe teada. Esimesena hakkas sellele probleemile tähelepanu pöörama läti päritolu Martins Pauls, kes pikkade aastate jooksul tegi hoolikalt ülestähendusi triinude sugupuude kohta ning kinnitas samuti juba ammu trinoloogide seas levinud teadmist, et triinud mitte kunagi ei ristu omavahel vaid ainult Homo sapiens sapiens´iga . Martins Pauls asus uurima kuivõrd palju saavad Homo sapiens ja Trinus sp. järglasteks triine ja kui palju inimesi. Ent milliseid isendeid ta ka ei uurinud, mitte kunagi ei olnud saadud järglaste seas triine. Seevastu järgmises põlvkonnas esinesid triinud juba üpris regulaarselt. Selle põhjal formuleeris ta 1942. aastal Triinugeneetika esimese seaduse:
Triinugeneetika esimene seadus
Trinus sp. ja Homo sapiens sapiens´i ristamisel F1 põlvkonnas triine ei saada. Edasi asus ta uurima, milliste seaduspärasustega triinude geenid kanduvad edasi F2 põlvkonna järglastele. Üldiselt näisid talle kõik saadud andmed üpriski segased ja juhuslikud ent ometi jõudis ta mõningatele järeldustele, mis küll olid saadud vaid statistikale toetudes ent osutusid siiski piisaval määral tõeks. Ta formuleeris järgnevad seadused:
Triinugeneetika teine seadus
F2 põlvkonnas sünnib triine suhtes 1:3. Täpsemalt on lahknemine F1 põlvkonnas 1:2:1 (ehk siis vastavalt triinud:naised:mehed).
Triinugeneetika kolmas seadus.
Väljaspool F1 põlvkonda võivad triinugeenid päranduda varjatult vaid naiste kaudu. F1 põlvkonnas aga ka meeste kaudu.
Ja olgugi, et need seadused tõesti kehtisid ei viinud need teadust kuigivõrd edasi, sest keegi ei osanud neid põhjendada ega selgitada. Pealegi oli vahepeal püstitatud tri-iini ehk kolm korda parandatud aminohappe hüpotees ja järjest rohkem trinolooge, kel enamasti polnud molekulaarbioloogiast õrna aimugi, hakkasid uskuma seda fantastilist teooriat.
Mida siis kujutas endast tri-iini hüpotees?
1952 oli IFRP´sse astunud parandamatu „laborirott” Mathew Jackson, ja asunud kohe tegelema triinude uurimisega molekulaartasemel. Ühes oma katsete seerias, kus uuris triinude valgulist struktuuri, eraldas ta triinurakust valgu, mille lagundas aminohapeteks ja lasi viimased seejärel läbi elektroforeesi. Lisaks tavaliste aminohapete tuttavale pildile vaatas talle vastu midagi, mis näis olevat kolm korda suurem. Ta kordas eksperimenti mitmeid kordi ja sai ikka ja jälle sama tulemuse - üks aminohape oli kolm korda suurem ning vähe sellest, teda paistis olevat ka kolm korda rohkem kui teisi! Nii jõudiski ta järeldusele, et triinude ilu ja kõige muu saladus seisneb selles aminohappes, mille ta nimetas tri-iiniks . Kui ta 1955. aastal oma uurimused avaldas oli trinoloogia jõudnud juba sellisesse faasi, et oldi valmis uskuma ükskõik mida, peaasi et see kasvõi mingil moel triinude erilist olemust teaduslikult seletaks - tri-iini teooria oli selleks rohkem kui suurepärane. Ning Mathew Jackson oli peagi tehtud mees. Sellegipoolest ei hakanud ükski trinoloog seeläbi rohkem huvituma molekulaartrinoloogia võimalustest - nad olid lihtsalt saanud kaelast ühe vastiku probleemi ning võisid nüüd rahus oma endise tööga edasi tegelda.
Kuid siiski leidusid kaks isepäist meest kellele me tänapäeval suure tänu võlgneme.
Esimene neist oli sakslane Adolf Bergenmeister, kes hakkas tegema ridamisi suuri avastusi kohe, kui asus uurima triinude genoomi.(Viimasest tuleb täpsemalt juttu allpool.)
Teine mees, Scott Fredricson, olles ise Mathew Jacksoni õpilane, lükkas aga ümber tri-iini hüpoteesi. Asi oli nimelt selles, et Jackson oli väga kokkuhoidlik inimene ja ehkki ta püüdis teha oma katsed äärmiselt kompetentselt, katsus ta siiski kokku hoida igal võimalikul juhul. Üks asi, mille arvelt tal juba aastaid oli õnnestunud kokku hoida, oli pipetiotsikute hankimine. Juba pikka aega oli ta püsikliendiks ühele mehele, kes müüs talle neid pea kümme korda odavamalt. Friedricson otsustas aga kontrollida, miks too mees neid nii odavalt müüs. Ta analüüsis otsikute kontsistentsi ja eraldas sealt kerge vaevaga ühendi, millel oli omadus siduda endaga 2 aminohapet. Rohkemat polnudki tarvis - veel mõned kinnitavad katsed ja tri-iin võis vajuda igaveseks ajaloo prügikasti.
Nüüd aga jätame ajaloo ja räägime sellest, mida trinoloogia tänaseks teada on saanud...
Triinude kromosoomistik ja trinosoomia
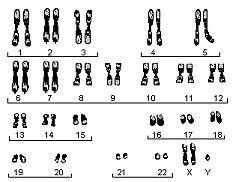
Joonis 7. Inimese karüogramm.
Nagu me kõik hästi teame, on inimesel 46 kromosoomi - 22 paari homoloogilisi ja 1 paar sugukromosoome, kusjuures naistel XX ja meestel XY. (Inimese karüogramm on toodud parempoolsel joonisel.) 22 homoloogilise kromosoomi osas on triinude ja inimeste karüogramm täiesti identne, kuid sugukromosoomidega paistab midagi totaalselt korrast ära olevat. Kui me võtame võrdluseks naise, kellega triinud väliselt vägagi sarnanevad, siis ilmneb et triinul on üks X-kromosoom kasvanud topelt nii pikaks, kui ta naisel oli, teise X-i asemel on aga (nii uskumatu kui see ka ei tundu) Y-kromosoom.(Triinu karüogramm on toodud alumisel joonisel.)
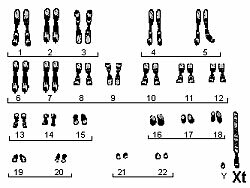
Joonis 8. Triinu karüogramm.
Selles suures X-kromosoomis ehk trinosoomis (Xt) asubki võti triinu saladuse avastamiseks. Xt üks osa on homoloogne naise X-kromosoomiga, ülejäänu aga kodeerib (nii nagu tänapäeva molekulaartrinoloogid öelda armastavad) trinosoomiat ehk triinuks olemist. Milleks on triinudel aga Y-kromosoom on siiani jäänud küllaltki selgusetuks . Samuti on segane see, miks avaldub trinosoomia vaid juhul kui on Xt Y ja miks ei avaldu trinosoomia kunagi F1 põlvkonnas? Kui me ristame triinu (Xt X) isase Homo sapiens´iga (X Y), siis saame me F1 põlvkonnas järgmised genotüübid:
- Xt X - naine, kes on trinosoomia varjatud kandja.
- Xt Y - peaks olema triin aga on mees, kes on trinosoomia varjatud kandja.
- X Y - tavaline mees.
- Y Y - sellise kombinatsiooni tekkel katkeb organismi areng juba peale paari esimest mitoosi.
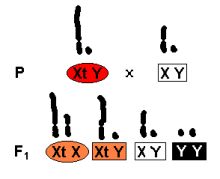
Joonis 9. Ristamisskeem.
Kui me järgnevalt toimetame ristamisi nii F1 põlvkonna siseselt kui ka puhaste (trinosoomiavabade) liinidega saame me mitmeid erinevaid kombinatsioone, milledest trinoloogia seisukohalt on eriti oluline DTS ehk topelt-trinosoomia: Xt Xt - kus isend on fenotüübilt naine, ent kannab varjatult mõlemas kromosoomis trinosoomiat määravaid geene. Sellise naise ristamisel on järglaste seas võrdselt nii triine kui trinosoomiat varjatult kandvaid naisi. Veelgi tähtsam on aga aspekt, et sellise kombinatsiooni puhul toimub trinosoomide vahel krossingover, mis liigisisesevarieeruvuse säilitamise seisukohalt mängib vägagi tähtsat rolli. Seda, milliseid kombinatsioone võib erinevatel ristamistel tekkida, võite te juba iseseisvalt edasi mõelda; meie suundume siinkohal uurima eelpoolnimetatud protsesside sügavamaid ja peenemaid tagamaid.
Trinosoomi osad ja X-kromosoomi kompensatsioon
Trinosoom jaotub kolmeks funktsionaalseks osaks:
- X-kromosoomiga homoloogiline osa.
- Trinosomaalseid geene kandev osa.
- X-kromosoomi kompenseeriv osa.
Esimeses osas paiknevad samad geenid, mis naise X-kromosoomis; millest tulenevalt on kombinatsioonis Xt X X-kromosoomi alleelid viimasega kompenseeritud ning ei teki sellist situatsiooni nagu meeste puhul, kus X-kromosoomi alleelid on „üksi jäetud” ja avalduvad olenemata sellest, kas on retsessiivsed, dominantsed...
Teises osas paiknevad kõik vaid triinudele omaseid tunnuseid määravad geenid, millest mitmetel on tähtis roll trinosoomia avaldumise regulatsioonis.
Kolmandas osas asuvad taas samad geenid mis X-kromosoomis, ainult et selle vahega, et neid X-kromosoomi geene, mis on olemas Y-kromosoomis, seal ei leidu. Põhjus on lihtne. Kui tekib kombinatsioon Xt Y (st. tekib trinosoomia), siis võetakse Xt 3. osa kui Xt 1. osale komplementaarset ahelat (ehk siis kui X-kromosoomi); need Xt 1. osa geenid millele Xt 3. osas „paarilist” ei leidu, leiavad endale homoloogi Y kromosoomi vastavatest osadest.
Trinosoomia avaldumist määravad geenid
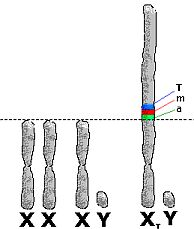
Joonis 10. Triinu kromosoomid.
Üks tähtsamaid ja esimesena avastatud trinosomaalseid geene on Trin-A, mille järgi sünteesitakse samanimelist valku, mis osaleb otseselt trinosoomia tekkes ja kujunemises.
Trin-A ise koosneb kolmest põhipiirkonnast - Trin-Az, promootor ja Trin-Aa. Trin-Az kodeerib valku UnZ, mis mingil kombel, mis veel tõnaseks teada pole, kontrollib, Y-kromosoomi olemasolu. Kui ta viimast ei avasta, siis seondub ta geeni Trin-M promootoralaga ja aktiveerib viimase transkriptsiooni. Kui ta aga Y kromosoomi olemasolu tuvastab, seostub ta sellesama Trin-A promootorpiirkonnaga, ja toimib seal samuti kui aktivaator ning laseb toimuda transkriptsioonil ja valgu Trin-A sünteesil. Valku Trin-A sünteesitakse ohtralt ja see osaleb pea kõikides komplitseeritud protsessides, mis peavad tagama valkude sünteesi trinosomaatilistelt geenidelt.
Trin-A ekspressiooni regulatsiooni kohta vaata ka videot.
Teise geeni, Trin-M, mida ka juba mainitud, pealt toimub transkriptsioon, kui tema promootoralale on kinnitunud valk UnZ. Sellisel juhul sünteesitakse valk Stop16, mis haakub tihedalt kromosoomi trinosomaalseid geene kandva osa ümber ega lase enamikku raku protsesse DNA-le ligi.
Kolmas geen, mis funktsioneerib sarnaselt eelmisega, kannab nime Trin-T. Tema avaldumine on ühelt poolt reguleeritud Trin-M´i toimega, ent teiselt poolt seotud mingite senitundmatute mehhanismide kaudu sellega, kas isendi ema on triin või mitte (vaata triinugeneetika esimest seadust). Kui Trin-T avaldub, sünteesitakse valk Stop15, mis haakub nii trinosoomi Trinosomaalseid geene kandva kui X-kromosoomi kompenseeriv osaga ja pärsib seeläbi vastavate alleelide avaldumise. Tulemuseks on isend, kes on fenotüübilt mees, kuid kannab varjatud trinosoomiat.
Siinkohal me lõpetame oma retke molekolaartrinoloogia telgitagustesse ja astume vastu praktilisele trinoloogiale.
Kirjutatud 1. novembril 2000.